





서 론
평균 수명의 증가로 노인 인구가 급증하면서 파킨슨병, 알츠하이머병, 루게릭병, 다발성 경화증 등의 퇴행성 뇌질환이 심각한 사회 문제로 대두되고 있다. 퇴행성 뇌질환의 정확한 발병 원인은 아직 불분명 하지만, 생체 대사과정에서 과도하게 축적된 활성산소가 DNA 손상, 단백질 산화, 지질과산화 증가, 신경세포 사멸 등을 유발한다고 알려졌다(Oh et al., 2016; Reddy et al., 2009). 현재까지 근본적인 퇴행성 뇌질환 치료제는 없는 실정으로 예방 및 개선을 목적으로 염증 억제, 세포사멸 억제, 인지능력 향상 활성이 높은 기능성물질 탐색 및 소재 개발 연구가 활발히 진행되고 있다.
프로바이오틱스(probiotics)는 숙주의 건강에 유익한 영향을 주는 살아있는 미생물로, 장내 미생물 균형 조절, 유해한 미생물의 침입과 정착 억제, 면역 반응 조절, 소화기능 개선, 아토피 개선, 신경염증 감소, 인지능력 향상 등 다양한 메커니즘을 통해 유익한 효과를 발휘한다(Isolauri et al., 2004, Williams et al., 2010). 일부 Lactobacillus와 Bifidobacterium 속 균주가 장-뇌축(gut- brain axis)에 영향을 주는 신경전달물질인 GABA와 serotonin을 분비하여 뇌 건강과 인지 기능에 유익한 영향을 주는 것으로 보고되었다(Dinan et al., 2013; Roshchina, 2010). GABA는 중추신경계에서 과도한 신경 흥분을 억제하여 이완을 촉진하고 스트레스 감소와 수면의 질을 향상하는데 도움을 주며, serotonin은 기분, 식욕, 수면 등을 조절하여 스트레스와 불안감을 감소하는 역할을 한다(Nam et al., 2009; Park, 2024). Lactobacillus plantarum MTCC1325와 Lactobacillus pentosus var. plantarum C29 균주의 섭취가 알츠하이머 유발 동물실험에서 기억 장애를 완화시키는 것으로 보고되었다(Nimgampalle and Kuna, 2017; Woo et al., 2014). 최근에는 열처리 유산균 사균체가 산화 스트레스가 유발된 신경세포에서 세포사멸을 억제하고 뇌유래신경영양인자(BDNF)의 발현을 촉진한다고 알려졌다(Cheon et al., 2021). 이러한 연구 결과들은 프로바이오틱스 생균, 사균체, 대사산물 등 미생물 소재가 불안, 우울증, 퇴행성 뇌질환 등 신경학적 질환의 예방, 개선 및 치료를 위한 소재로 활용될 가능성을 제시하고 있다.
콩은 전 세계적으로 중요한 식량 자원 중 하나로, 단백질, 지방, 섬유질, 비타민 B군, 미네랄 등 영양 성분이 풍부하며(Ferreira et al., 2018), isoflavone, anthocyanin, saponin 등 생리활성 물질을 함유하고 있어 항산화, 항염, 항암 활성을 나타낸다고 알려졌다(Cho et al., 2007). 특히, 검은콩은 anthocyanin과 isoflavone이 풍부하여 산화적 스트레스 억제 활성이 높고 신경세포 보호 효과가 있다고 보고되었다(Im et al., 2006). 검은콩 껍질에 존재하는 총 anthocyanin의 약 70%를 차지하는 cyanidin- 3-glucoside 성분이 산화 스트레스가 유도된 PC12 신경세포의 세포사멸에 대한 보호 효과를 나타냈으며(Kwak et al., 2013), 대두의 isoflavone이 amyloid β peptide(Aβ)에 의해 유도된 학습과 기억장애를 개선하였다고 보고되었다(Devi et al., 2017). 하지만 현재까지 검은콩과 유산균을 활용하여 정신 건강에 도움이 되는 기능성 발효 소재의 개발에 관한 연구는 거의 전무한 실정이다.
최근 프로바이오틱스 제품 시장이 빠르게 성장하고 있지만, 생균 섭취로 인한 알레르기, 설사, 가스 발생, 복통, 두통 등의 부작용 사례가 증가하고 있으며 일부 면역억제 환자에게서 패혈증과 균혈증이 보고되는 등 심각한 안전성 문제가 제기되고 있다. 이러한 상황에서 최근에는 프로바이오틱스 균주의 사균체와 발효물이 항산화, 항염, 면역 조절, 인지능력 향상 등 생균과 유사한 유용한 효과를 나타낸다고 밝혀져 안전성이 높은 대체제로 주목받고 있다(Nataraj et al., 2020; Siciliano et al., 2021). 현재까지 국내의 사균체 연구는 열처리한 유산균 사균체를 이용한 소수의 연구만이 보고되었으며, 사균체를 함유한 발효물의 연구는 거의 이루어지지 않았다. 사균체 함유 검은콩 발효물은 불안, 우울증, 퇴행성 뇌질환 등 신경학적 질환의 개선을 위한 건강식품 소재로 활용될 수 있는 잠재력을 가지고 있어 이에 대한 관심과 집중적인 연구가 필요하다.
본 연구자들은 선행 연구에서 김치로부터 프로바이오틱스 특성이 우수한 2종의 유산균주(L. plantarum SMF470, L. plantarum SMF796)를 선별하였으며, 이들의 열처리 사균체가 항산화 및 항염 활성이 우수하다는 것을 확인하였다(Lee et al., 2022; You et al., 2022). 본 연구에서는 L. plantarum SMF470과 L. plantarum SMF796 균주를 스타터로 이용하여 사균체를 함유한 열처리 검은콩 발효물을 제조하고, H2O2에 의한 산화적 스트레스와 MPP+에 의해 유도된 신경세포 손상에 대한 보호 활성을 측정함으로써 사균체 함유 발효물을 활용한 퇴행성 뇌질환 개선 연구의 기초자료를 확보하고자 하였다.
재료 및 방법
선행 연구를 통해 김치로부터 프로바이오틱스 특성이 우수한 2종의 유산균주(L. plantarum SMF470, L. plantarum SMF796)를 선별하였으며, 본 연구에서 검은콩 발효를 위한 스타터로 사용하였다. 상업용 프로바이오틱스인 Lacticaseibacillus rhamnosus GG(Korean Collection for Type Cultures, KCTC 5033) 균주는 생물자원센터(Daejeon, Korea)에서 분양받았다. 발효에 사용한 검은콩 분말은 그레인팜(Jangheung-gun, Korea)에서 구입하였다. 인간 결장 유래 HT-29 세포(KCLB30038)와 인간 신경모세포종 SH-SY5Y 세포(KCLB22266)는 한국세포주은행(Seoul, Korea)에서 분양받았다.
검은콩 분말 5%(w/v)를 증류수에 첨가하고 121℃에서 15분 동안 열수 추출하여 0.45 μm syringe filter(Whatman, England)로 여과 후 검은콩 추출물을 생성하였다. 검은콩 추출물에 식용 protease인 Alcalase 2.4L(Novozyme, Bagsvaerd, Denmark)와 Flavourzyme 1000L(Novozyme, Bagsvaerd, Denmark)을 1%(w/v) 농도로 첨가하고 50℃에서 2시간 동안 반응시킨 후 100℃에서 5분간 가열처리하여 효소를 불활성화하였다. 이후 검은콩 추출물을 동결건조하여 분말화한 후 20~30℃에 보관하여 실험에 사용하였다. 동결건조된 검은콩 추출물 분말을 멸균 증류수에 5%(w/v) 첨가한 것을 유산균 발효물을 제조하기 위한 검은콩 배지로 사용하였다. Lactobacilli MRS broth(BD DifcoTM, USA)에 37℃, overnight 배양한 유산균 배양액(L. rhamnosus GG, L. plantarum SMF470, L. plantarum SMF796)을 검은콩 배지에 약 2×107 CFU/mL 수준이 되도록 2%(w/v) 접종한 후 37℃에서 24시간 발효를 진행하였다. 회수한 발효물을 121℃에서 15분간 멸균 처리하여 사균체를 함유한 열처리 발효물 시료로 사용하였다.
검은콩 발효 기간 동안 발효액의 생균수(CFU/mL)와 OD600nm, pH 변화를 측정하였다. 발효액 시료 100 μL에 멸균 생리식염수를 첨가하여 10배수로 희석하고, MRS agar에 도말한 후 37℃에서 48시간 동안 배양하고 콜로니 수를 계산하여 생균수(CFU/mL)를 측정하였으며, 균체량을 확인하기 위해 600 nm에서 흡광도(Biochrom Libra S22 UV/Vis Spectrophotometer, USA)를 측정하였다. pH는 pH meter(Seven CompactTM pH/Ion S220, Meltter-Toledo, Switzerland)를 이용하여 측정하였다.
DPPH(1,1-diphenyl 1-2-picrylhydrazyl) 라디칼 소거능은 Wang 등(2016)의 방법을 응용하여 측정하였다. 0.4 mM DPPH 용액 1 mL와 발효물 시료 1 mL를 혼합하여 암실에서 30분간 반응시킨 후 원심분리(13,000×g, 4℃, 10 min, CT15RE, Hitachi Chemical Co., Ltd., Japan)하여 상등액을 회수하였다. 상등액을 흡광도 517 nm에서 측정한 후 아래의 계산식에 따라 DPPH 라디칼 소거능(%)를 계산하였다. 대조구로 Phosphate buffered saline(PBS, Welgene, Korea) 용액을 사용하였다.
ABTS(2,2’-azino-bis 3-ethylbenzothiazoline-6-sulphonic acid) 라디칼 소거능은 Ji 등(2015)의 방법을 응용하여 측정하였다. 0.2 M sodium phosphate buffer(pH 7.4)에 7 mM ABTS와 5 mM potassium persulfate를 혼합하고 암실에서 16시간 이상 안정화시킨 후 734 nm에서 흡광도를 0.7로 조정하여 ABTS 시약을 제조하였다. ABTS 시약 1.35 mL와 발효물 시료 150 μL를 혼합하고 37℃에서 10분간 반응시킨 후 원심분리(13,000×g, 4℃, 10 min)하여 상등액을 회수하였다. 상등액의 흡광도를 734 nm에서 측정하고 아래 계산식에 따라 ABTS 라디칼 소거능(%)을 계산하였다. 대조구로 PBS 용액을 사용하였다.
총 폴리페놀 함량은 Ahn 등(2012)의 방법을 응용하여 측정하였다. 발효물 시료 0.5 mL를 1 N Folin & Ciocalteu's phenol reagent(St. Louis, MO, USA) 0.3 mL, 2% NaCO3 3 mL와 혼합하여 상온에서 30분간 반응시킨 후 720 nm에서 흡광도를 측정하였다. 표준물질로 gallic acid를 사용하여 표준곡선을 작성한 후 gallic acid equivalent(mg GAE/g)로 환산하여 시료의 총 폴리페놀 함량을 계산하였다.
HT-29 세포는 Roswell Park Memorial Institue 1640(RPMI, Welgene, Korea)배지를 사용하였으며, SH-SY5Y 세포는 Dulbecco’s modified Eagle’s medium(DMEM, Welgene, Gyeongsan, Korea)을 사용하였다. 각 배지에 10% Fetal Bovine Serum(FBS, GW Vitek, Seoul, Korea)과 1% penicillin-streptomycin solution(Cytiva, Marlborough, USA)을 첨가한 후 37℃, 5% CO2incubator(Thermo Fisher, Massachusetts, USA)에서 세포를 배양하였다.
MTT(methylthizol-2-yl-2,5-diphenyl tetrazoliumbromide) 방법에 따라 HT-29 세포에 대한 발효물 시료(undiluted solution, 1/2 희석시료, 1/5 희석시료, 1/10 희석시료)의 세포독성을 평가하였다(Chuah et al., 2019). 96-well plate에 HT-29 세포를 1×105 cell/well 수준으로 접종한 후 37℃, 5% CO2에서 24시간 배양하였다. 배양액으로부터 상등액을 제거한 다음 RPMI 180 μL와 발효물 20 μL를 첨가하여 24시간 배양한 후 well에 남아있는 발효물을 PBS 용액으로 두 번 세척하였다. 이후 RPMI 180 μL와 MTT 시약(5 mg/mL) 20 μL를 첨가하여 4시간 동안 반응하였다. 반응 후 생성된 formazan 침전물에 dimethyl sulfoxide(DMSO, Daejung, Siheung, Korea) 100 μL를 첨가하여 용해시킨 후 SPECTROstar Nano microplate reader(BMG labtech, Germany)로 570 nm에서 흡광도를 측정하였다. 생존율(%)은 아래 계산식에 따라 계산하였으며, 대조구로 PBS 용액을 사용하였다.
뇌장축을 통한 열처리 검은콩 발효물의 신경보호 효과를 평가하기 위하여, 열처리 추출물 또는 열처리 발효물을 인간 결장 유래 HT-29 세포와 배양하여 열처리 추출물 유래 조건배지(heat- treated black bean extracts-conditioned medium, BE-CM)와 열처리 발효물 유래 조건배지(heat-treated fermented black beans-conditioned medium, HFB-CM)를 제조하였다(Lim et al., 2020). HT-29 세포를 6-well plate에 1×105 cell/well 농도로 접종하여 48시간 동안 배양한 후, RPMI 1.8 mL와 열처리 추출물과 발효물 200 μL를 첨가하였으며, 최종 처리 농도를 1/10배 희석된 농도로 처리하여 24시간 동안 배양하였다. 이후 상등액을 회수하여 10분 동안 원심분리(13,000×g, 4℃)한 후 0.45 μm syringe filter로 여과하여 조건배지를 제조하였다. 조건배지 시료는 –80℃에서 보관하며 사용하였다.
MTT 방법에 따라 인간 신경모세포종 SH-SY5Y 세포에 대한 조건배지의 세포독성을 평가하였다(Lim et al., 2020). 96-well plate에 SH-SY5Y 세포를 1×105 cell/well 수준으로 접종한 후 24시간 동안 배양하였다. 상등액을 제거한 후 DMEM 180 μL와 조건배지 20 μL를 첨가하였으며, 최종 처리 농도를 1/10배 희석된 농도로 처리하여 24시간 동안 배양한 다음 PBS 용액으로 두 번 세척하였다. 이후 DMEM 180 μL와 MTT 시약(5 mg/mL) 20 μL를 첨가하여 4시간 동안 반응하였다. 반응 후 생성된 formazan 침전물에 DMSO 100 μL를 첨가하여 용해시킨 후 microplate reader를 이용하여 570 nm에서 흡광도를 측정하였다. 대조구는 PBS 용액을 처리하여 진행하였으며, 생존율(%)은 아래 계산식에 따라 계산하였다.
조건배지의 산화적 스트레스에 대한 신경세포 보호 효과 평가는 Park 등(2017)의 방법을 응용하여 실험하였다. 96-well plate에 SH-SY5Y 세포를 1×105 cell/well 수준으로 접종한 후 37℃, 5% CO2에서 24시간 동안 배양하였다. 이후 상등액을 제거하고 DMEM 100 μL와 조건배지 80 μL를 첨가하여 4시간 배양한 후 20 μL의 H2O2로 20시간 처리하였다. PBS로 세포를 2번 세척한 후 DMEM 180 μL와 5 mg/mL 농도의 MTT 용액 20 μL를 첨가하여 4시간 동안 반응하였다. 상등액을 제거하고 생성된 formazan 침전물을 DMSO 100 μL로 용해시킨 후 microplate reader를 이용하여 570 nm에서 흡광도를 측정하였다. 대조구는 PBS 용액을 처리하여 진행하였으며, 생존율(%)은 아래 계산식에 따라 계산하였다.
Quantitative reverse transcription-polymerase chain reaction(qRT-PCR)을 이용하여 BDNF, COX-2, iNOS, BAX, BCL-2 유전자의 mRNA 발현을 측정하였다(Cheon et al., 2020). SH-SY5Y 세포를 1×105 cell/well 농도로 6-well plate에 접종하고 37℃, 5% CO2에서 24시간 배양하였다. 배양액으로부터 상등액을 제거하고 DMEM 1 mL와 조건배지 800 μL를 첨가하여 4시간 처리하였다. 이후 1 mM MPP+가 첨가된 DMEM 배지 200 μL를 첨가하고 20시간 반응하였다. 반응 종료 후 PBS 용액으로 두 번 세척하고 세포를 회수하여 PCR 분석에 사용하였다. Total RNA는 RNeasy mini kit(Qiagen, Hilden, Germany)를 이용하여 추출하였고, NanoDrop(biochrom, Cambridge, USA)을 이용하여 농도를 정량하였다. 정량한 RNA는 cDNA synthesis kit(TAKARA, Shigaken, Japan)를 이용하여 합성하였으며, StepOnePlusTM Real-time PCR System(Thermo Fisher, Massachusetts, USA)을 이용하여 유전자 발현을 측정하였다. Primer(Cosmogenetech, Seoul, Korea) 2 μL, cDNA 2 μL, SYBR green(Cosmogenetech, Seoul, Korea) 10 μL, 증류수 6 μL를 혼합하여 PCR을 진행하였다. House keeping gene으로 GAPDH를 사용하였다. PCR은 95℃에서 15초(denaturation), 60℃에서 60초(annealing/extension) 반응으로 40회 반복하여 진행하였으며, delta-delta Ct(ΔΔCt)방법을 이용하여 결과를 분석하였다. PCR primer는 코스모진텍(Seoul, Korea)에서 구매하여 사용하였다(Table 1).
| Primers1) | Sequences | References |
|---|---|---|
| BDNF | F:5‘-CAAACATCCGAGGACAAGGTGG-3’ | Cheon et al., 2020 |
| R:5‘-CTCATGGACATGTTTGCAGCATCT-3’ | ||
| BAX | F:5‘-GTGGTTGCCCTCTTCTACTTTGC-3’ | Cheon et al., 2020 |
| R:5‘-GCGGACTCCAGCCACAAAGATG-3’ | ||
| BCL-2 | F:5‘-CGGCTGAAGTCTCCATTAGC-3’ | |
| R:5‘-CCAGGGAAGTTCTGGTGTGT-3’ | ||
| iNOS | F:5‘-CCTCGCTCTGGAAAGACCAG-3’ | Cui et al., 2019 |
| R:5‘-CATGGGTTTTCCAGGCCTCT-3’ | ||
| COX-2 | F:5‘-AGCATCTACGGTTTGCTGTG-3’ | Ma et al., 2019 |
| R:5‘-CCTGTTTAAGCACATCGCAT-3’ | ||
| GAPDH | F:5‘-GAGTCAACGGATTTGGTCGT-3’ | Cheon et al., 2020 |
| R:5‘-GACAAGCTTCCCGTTCTCAG-3’ |
결과 및 고찰
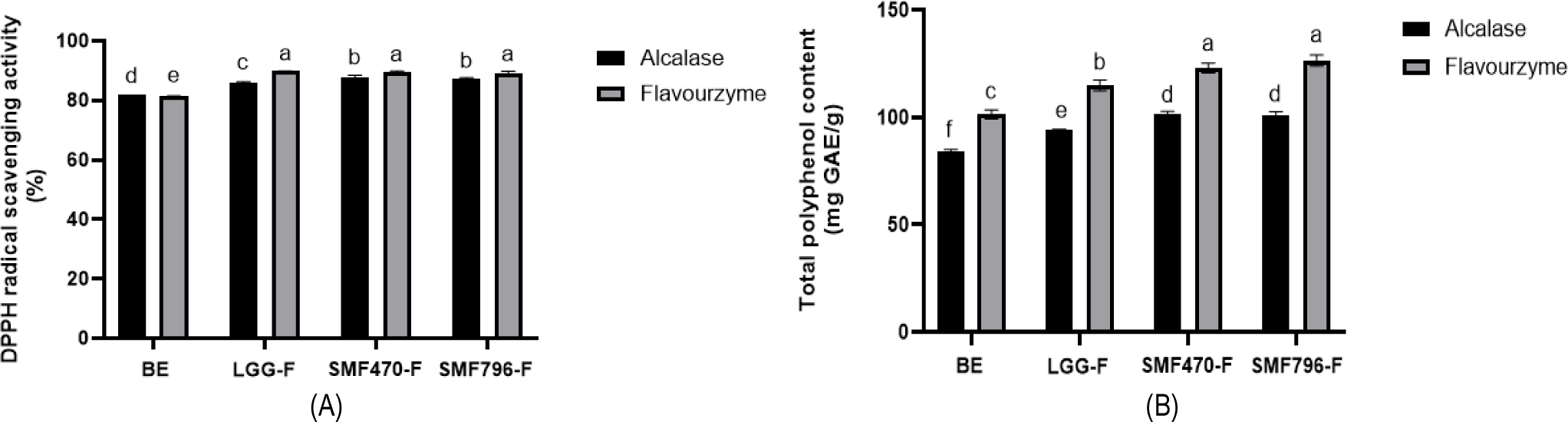
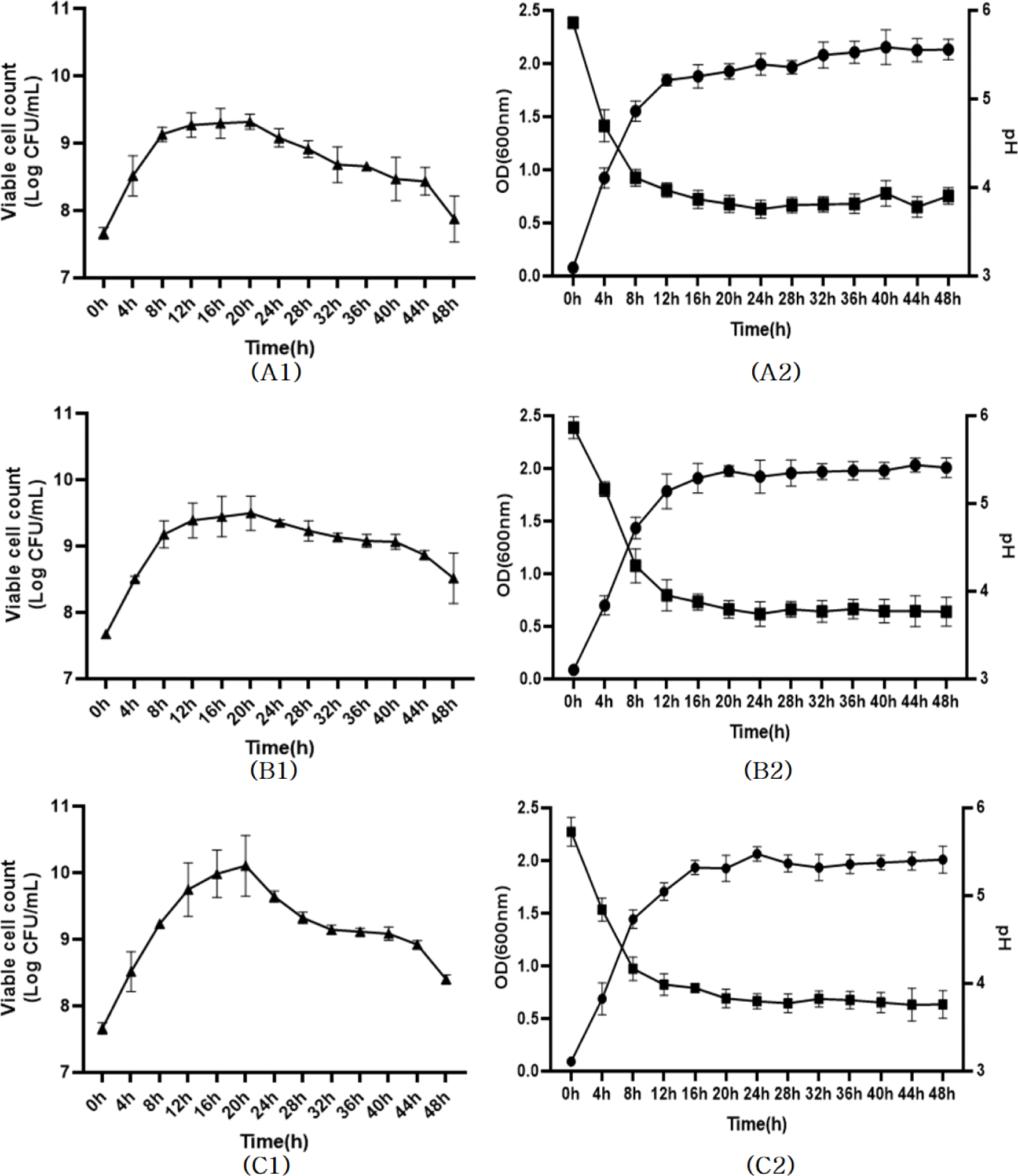
유산균 발효를 위한 검은콩 추출물 배지를 최적화하기 위하여 검은콩 농도, 열수 추출 온도와 시간, protease 처리 조건을 검토하였다. 검은콩 분말 5%(w/v)를 121℃에서 15분간 열수 추출한 후 산업용 protease인 Alcalase와 Flavourzyme으로 처리하여 발효 후 항산화 활성을 측정한 결과, Flavourzyme으로 처리한 발효물에서 유산균의 생육 및 항산화 활성이 우수한 것이 확인되어(Fig. 1), 유산균 발효를 위한 검은콩 추출물 배지 조건으로 Flavourzyme을 처리하는 것으로 선정한 후 실험을 진행하였다. 선행 연구를 통해서 김치로부터 프로바이오틱스 활성이 우수한 2종의 유산균주(L. plantarum SMF470, L. plantarum SMF796)를 선별하였으며, 이들의 열처리 사균체가 항산화 및 항염 활성이 우수하다는 것이 확인되었다(Lee et al., 2022; You et al., 2022). 본 연구에서는 유산균체 함량이 높은 검은콩 발효물을 제조하기 위해 검은콩 추출물 배지에 선별된 유산균과 산업용 프로바이오틱스인 L. rhamnosus GG(LGG)를 각각 2%(v/v) 접종하고 48시간 동안 배양하면서 유산균의 생육특성을 분석하였다(Fig. 2). LGG, SMF470, SMF796 균주의 생균수(CFU/mL)는 20시간 발효 후에 각각 최대 9.28, 9.40, 9.76 Log CFU/mL까지 증가하였으나, 24시간 이후부터는 생육이 저해되어 48시간에는 생균수가 감소하는 경향을 나타내었다. 이는 산 생성량 증가, 낮은 pH, 영양원 부족 등의 이유로 유산균의 생육이 억제되었기 때문인 것으로 사료된다(Hwang et al., 2022). OD600nm값은 24시간 발효 후 각각 최대 2.08, 2.06, 2.08까지 증가하였으며, 48시간까지 일정하게 유지되었다. 모든 발효물의 pH는 24시간 발효 후 3.83-3.84까지 감소하였으며, 이후 48시간까지 일정하게 유지되었다. 따라서 유산균 사균체를 함유한 발효물을 제조하기 위하여 24시간까지 발효한 검은콩 발효물을 선정하고 열처리 공정을 진행하였다.
검은콩 발효물을 121℃(15분)에서 열처리하여 유산균 사균체를 함유한 열처리 검은콩 발효물을 제조하고 DPPH 라디칼 소거능, ABTS 라디칼 소거능, 총 폴리페놀 함량을 측정한 결과는 Table 2와 같다. 열처리한 LGG 발효물(LGG-F), SMF470 발효물(SMF470-F), SMF796 발효물(SMF796-F)의 DPPH 라디칼 소거능은 각각 63.44%, 62.02%, 63.29%로, 대조구인 열처리 검은콩 추출물(black bean extract, BE)의 55.57%보다 유의적으로 높은 항산화 활성을 나타내었다(p<0.05). ABTS 라디칼 소거능에서도 LGG-F, SMF470-F, SMF796-F는 각각 81.25%, 81.73%, 81.37%의 소거능을 나타내어, BE(75.77%)와 비교하여 유의적으로 높은 항산화 활성을 보였다(p<0.05). 발효물의 총 폴리페놀 함량은 155.98-156.09 GAE mg/g으로 BE(118.74 GAE mg/g)보다 유의적으로 높은 함량을 나타내었다(p<0.05). 열처리한 검은콩 발효물에서 항산화 활성이 증가되는 것은 유산균 사균체 성분의 용출과 유산균에 의해 검은콩의 생리활성 성분인 isoflavone과 anthocyanin이 비배당체로 전환되었기 때문인 것으로 사료된다. 사균체의 생리활성 성분으로는 exopolysaccharide, peptidoglycan, lipoprotein, teichoic acid, cell wall protein 등이 알려졌으며(Akter et al., 2020), Kang 등(2011)은 유산균 사균체 첨가에 의해 발효유의 DPPH와 ABTS 라디칼 소거능이 농도 의존적으로 증가되었다고 보고하였다. Hwang 등(2018)은 분말 두유의 유산균 발효에 의해 DPPH와 ABTS 라디칼 소거능이 증가하였으며 이는 비배당체 isoflavone인 daidzein과 genistein 함량의 증가에 기인한다고 보고하였다.
a-b Values with different superscript letters in the same column are significantly different(p<0.05). BE (control); heat-treated black bean extract, LGG-F, SMF470-F, and SMF796-F; heat-treated fermented black bean products by LGG, SMF470, and SMF796, respectively. LGG; L. rhamnosus GG, SMF470; L. plantarum SMF470, SMF796; L. plantarum SMF796.
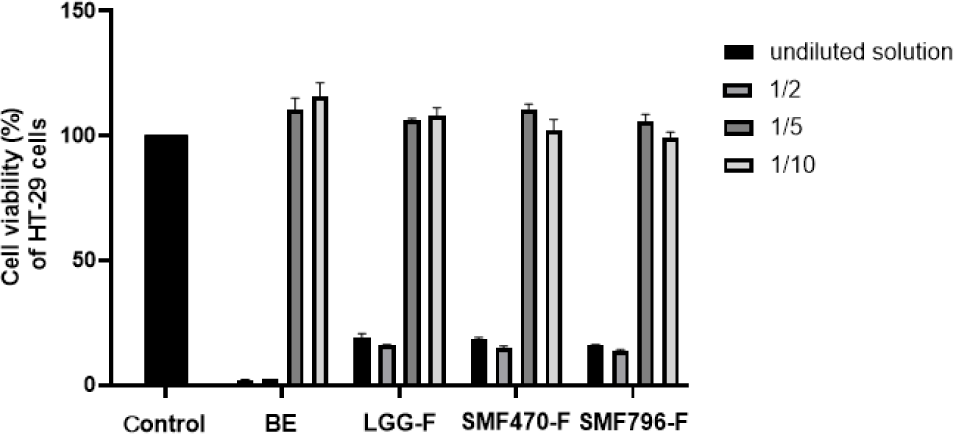
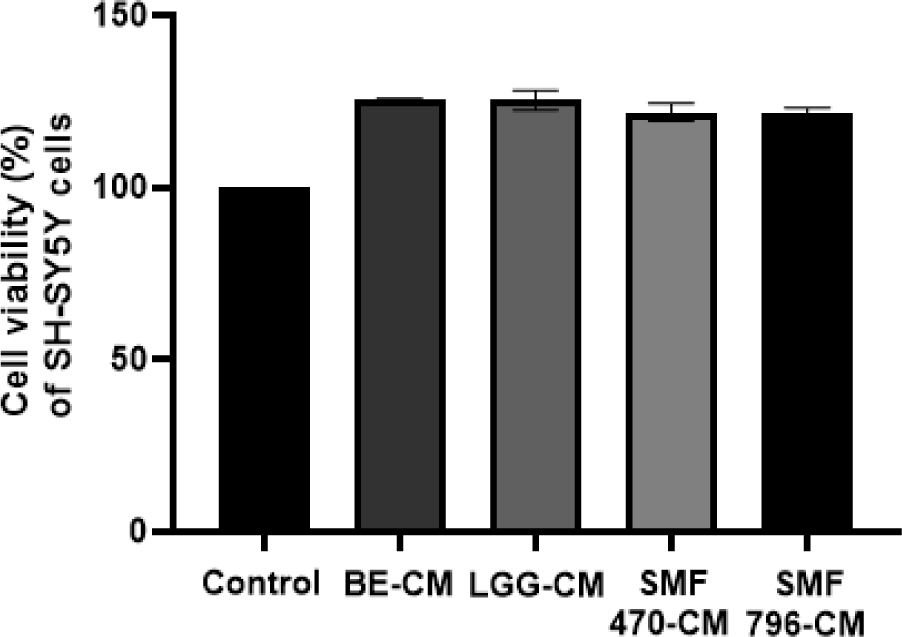
열처리 검은콩 발효물의 신경보호 효과를 평가하기 위하여, 발효물을 장세포(HT-29)에 처리하여 조건배지(CM)를 제조하고, MTT assay를 이용하여 세포독성을 평가하였다. MTT assay는 살아있는 세포가 미토콘드리아 탈수소효소에 의해 노란색의 MTT tetrazolium을 환원시켜 자주색의 formazan 결정을 형성하는 원리에 근거하며, 형성된 formazan 결정을 용해하여 흡광도를 측정하면 세포의 생존율을 정량적으로 평가할 수 있다(Carmichael et al., 1987). HT-29 세포에 열처리 검은콩 발효물을 농도별(undiluted solution, 1/2 희석시료, 1/5 희석시료, 1/10배 희석시료)로 처리하여 세포독성을 측정한 결과, 1/5배와 1/10배 희석한 추출물과 발효물 시료에 대하여 100% 이상의 생존율을 나타내었다(Fig. 3). 1/5배 희석한 추출물과 발효물에서 세포 독성이 나타나지 않는 것이 확인되어, HT-29 세포에 처리하여 조건배지를 제조하였다. SH-SY5Y 세포에 검은콩 추출물과 발효물로 제조한 조건배지를 처리하여 세포독성을 측정한 결과, 모두 100% 이상의 생존율을 보이는 것이 확인되었다(Fig. 4). 따라서 본 연구에서 사용한 열처리 발효물의 조건배지는 SH-SY5Y 세포에 독성을 나타내지 않아 신경세포 실험에 사용하기 적합하다고 판단된다.
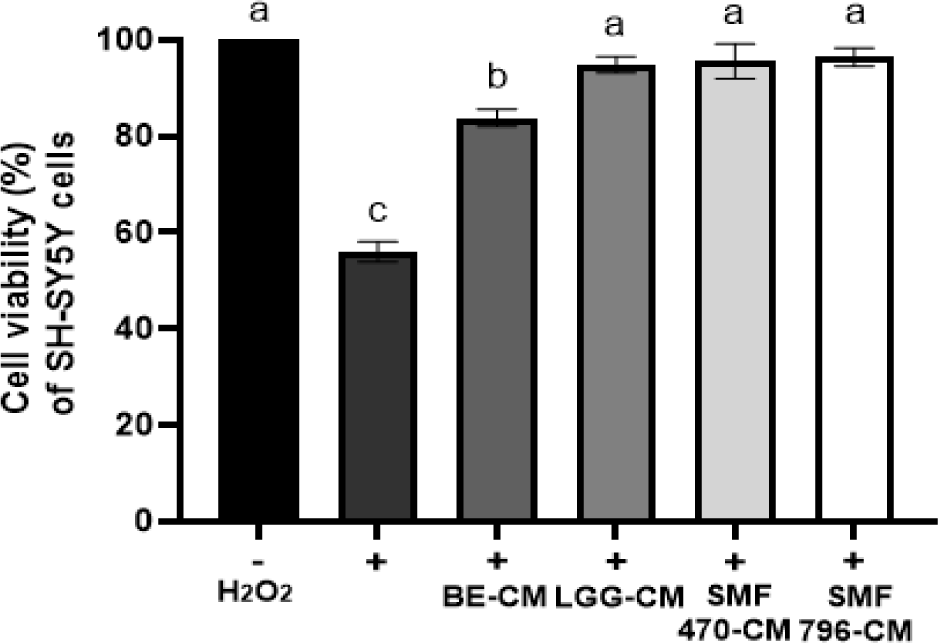
뇌조직은 고농도의 다중불포화지방산으로 구성되어 있으며, 다른 조직에 비해 산소 소모량이 높은 반면 낮은 항산화 시스템을 갖고 있어 산화적 스트레스로 인한 손상으로부터 취약하다(Omodeo-Sale et al., 1997). 산화적 스트레스는 DNA 손상, 단백질 산화, 지질과산화물 생성을 통하여 신경세포 손상 및 세포 사멸을 유발하므로 퇴행성 뇌질환의 주요 원인으로 보고되고 있다(Lee et al., 2018). H2O2에 의해 산화적 스트레스가 유발된 SH-SY5Y 세포에 추출물 유래 조건배지(BE-CM)와 발효물 유래 조건배지(HFB-CM)를 처리하고 세포 생존율을 측정한 결과는 Fig. 5와 같다. H2O2 비처리군(-)에서 SH-SY5Y 세포는 100%의 생존율을 보인 반면, 250 μM의 H2O2처리에 의해 50%에 가까운 세포 생존율을 나타내어, 산화적 스트레스 평가를 위한 H2O2처리 농도로 선정하였다. HFB-CM (SMF470-CM, SMF796-CM, LGG-CM) 처리군에서 세포 생존율이 94.83-96.40%를 나타내어, H2O2단독처리군(+)의 55.99%에 비해 약 38-40% 생존율이 높은 것을 확인할 수 있었다. BE-CM 처리군도 83.79%의 높은 세포 생존율을 보였으나, HFB-CM 처리군보다는 생존율이 유의적으로 낮은 것으로 나타났다(p<0.05). 본 연구결과 열처리 검은콩 추출물 또는 열처리 검은콩 발효물은 H2O2에 의해 산화적 스트레스가 유도된 신경세포를 보호하는 효과를 나타내는 것을 알 수 있었으며, 유산균 발효에 의해 보호 효과가 증가되는 것으로 확인되었다. Lee 등(2021)은 프로바이오틱 요구르트의 조건배지(PY-CM) 처리에 의해 H2O2에 의해 산화적 스트레스가 유도된 SH-SY5Y 세포의 생존율이 115% 증가되었다고 보고하여, 산화적 스트레스에 대한 PY-CM의 신경세포 보호 효과를 입증한 바 있다.
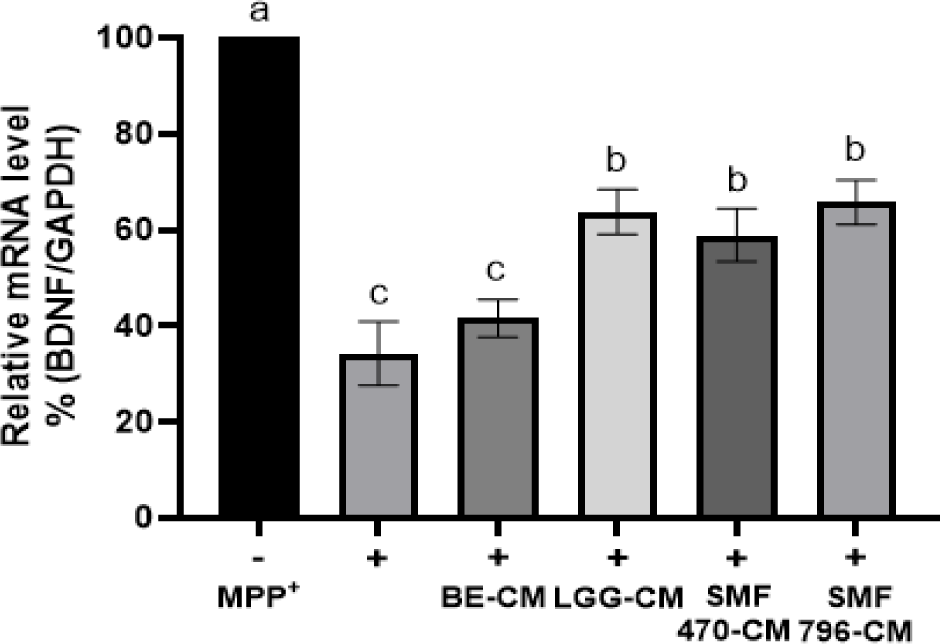
MPP+는 도파민 수송체를 통해 신경세포 내로 침투하여 미토콘드리아에서 산화적 스트레스와 세포 사멸을 유발하는 신호를 전달함으로써 파킨슨병 유사 증후군을 유도하는 것으로 알려져 있다(Eberhardt and Schulz, 2003; Przedborski and Jackson- Lewis, 1998). 본 연구에서는 MPP+에 의해 유발된 신경독성에 대한 열처리 검은콩 발효물 유래 조건배지의 신경보호 효과를 평가하기 위해, MPP+를 처리한 SH-SY5Y 세포에 CM을 첨가한 후 qRT-PCR을 이용하여 BDNF, COX-2, iNOS, BAX, BCL-2 유전자의 발현 변화를 측정하였다.
BDNF는 신경세포의 증식과 생존에 필수적인 역할을 하는 뇌유래신경영양인자로 중추신경계 및 말초신경계에 다량 존재한다. BDNF는 신경세포의 증식과 생존 및 시냅스의 성장과 분화에 중요한 역할을 담당하며, 신경세포를 보호하고 인지 능력을 향상시키는 데 도움을 준다고 알려졌다(Colucci-D'Amato, 2020; Murray et al., 1994). MPP+를 처리한 SH-SY5Y 세포에 추출물 유래 조건배지(BE-CM)와 발효물 유래 조건배지(HFB-CM)를 첨가하고 BDNF 발현량을 측정하였다(Fig. 6). MPP+ 처리(+)에 의해 세포독성이 유도된 SH-SY5Y 세포에서 BDNF 발현이 현저히 감소되는 것을 확인할 수 있었으며, HFB-CM 처리에 의해 MPP+에 의해 감소된 BDNF 발현이 1.72-1.92배 유의적으로 증가되는 것으로 확인되었다(p<0.05). 한편, BE-CM은 HFB-CM과 비교하여 유의적으로 낮은 BDNF 발현 수준을 보였다(p<0.05). Lee 등(2018)은 Lactobacillus plantarum C29를 이용하여 발효한 탈지 대두를 쥐에게 경구 투여하였을 때, 인지 기능 저하를 완화시키고 BDNF의 발현을 증가시켜 알츠하이머 증상을 개선시킬 수 있다고 보고하였다.
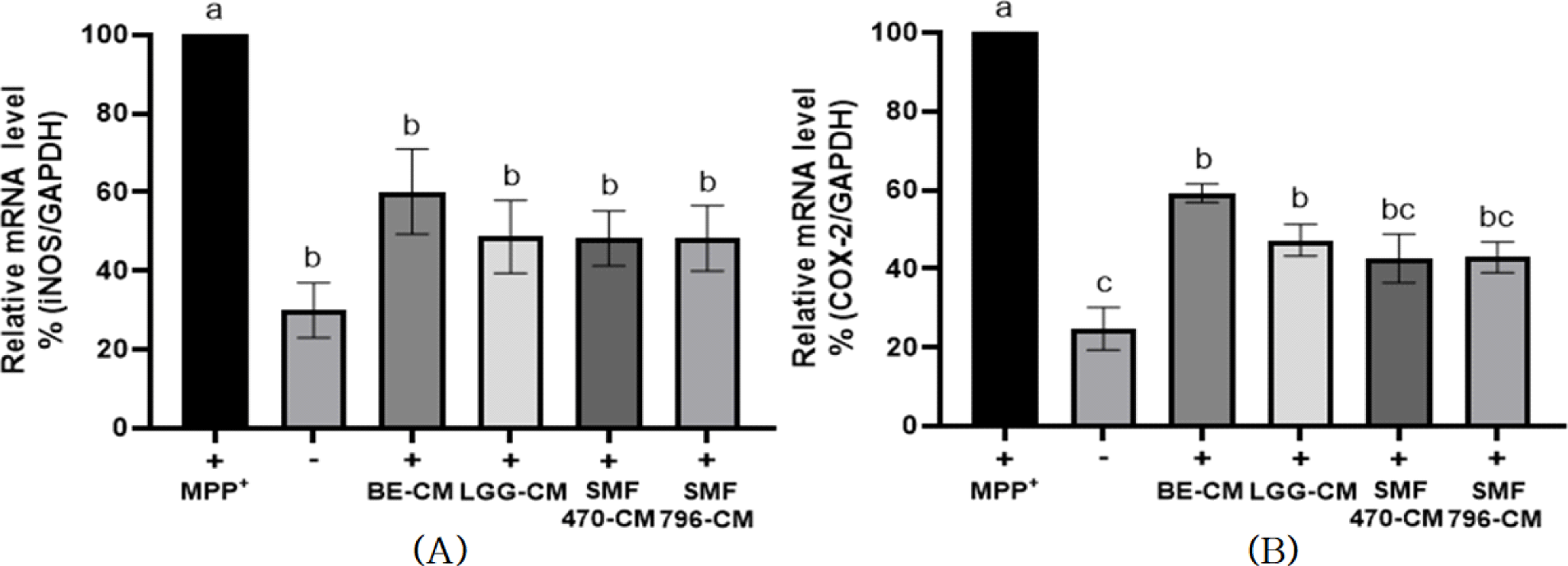
MPP+는 inducible nitric oxide synthase(iNOS)와 cyclooxygenase-2(COX-2)와 같은 염증성 매개인자들의 과도한 발현을 촉진하여 염증을 악화시킨다고 알려졌다(Cui et al., 2019; Wang et al., 2005). MPP+에 의해 신경독성이 유도된 SH-SY5Y 세포에 BE-CM과 HFB-CM을 처리하고 iNOS와 COX-2의 발현을 평가하였다(Fig. 7). MPP+ 단독처리군(+)과 비교하여, SMF470-CM, SMF796-CM, LGG-CM 처리군에서 COX-2의 발현이 각각 57.37%, 57.13%, 52.78% 억제되었으며, iNOS의 발현은 각각 51.74%, 51.73%, 51.36% 억제되었다. NO는 주로 iNOS의 효소작용에 의해 합성되며, 중추신경계에서 과도하게 생성된 NO는 염증, 조직 손상, 신경 손상을 유발하여 파킨슨병, 알츠하이머와 같은 퇴행성 뇌질환의 발병에 관여한다(Saha and Pahan, 2006). iNOS와 더불어 염증반응에 주요한 역할을 하는 효소인 COX-2는 통증과 발열을 유발하는 prostaglandin E2의 합성을 유도하며, 파킨슨병 동물모델에서 과도하게 발현된다(Wang et al., 2005). Lee 등(2023)은 발효차가 MPTP(1-Methyl-4-phenyl-1,2,3,6- tetrahydropyridine)에 의해 파킨슨병 유사 신경세포 손상이 유발된 생쥐에서 iNOS와 COX-2의 발현 수준을 현저히 감소시켜 도파민성 신경세포 손상을 억제한다고 보고하였다.
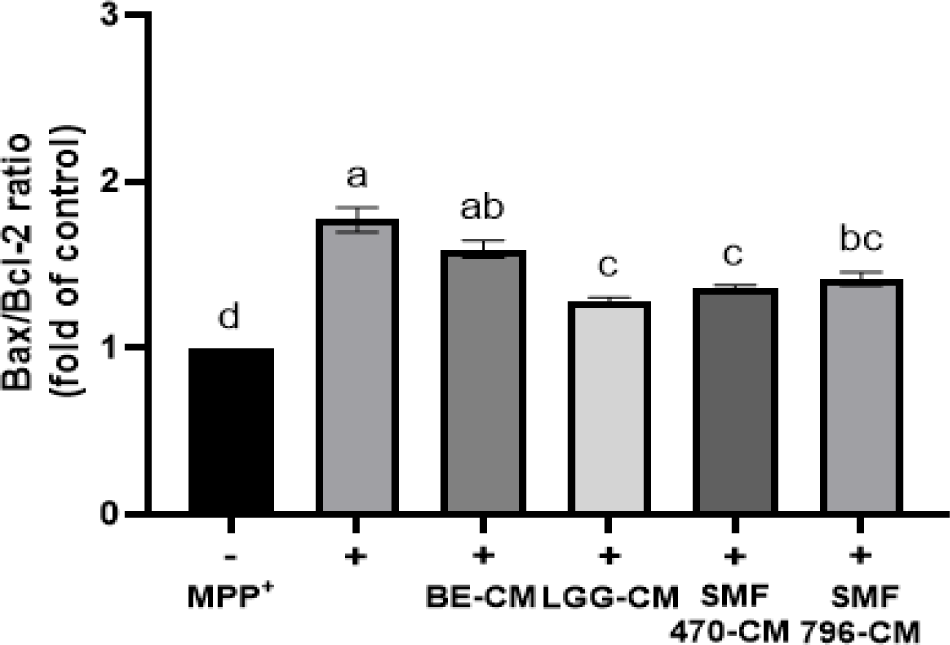
Bax와 Bcl-2는 미토콘드리아에서 세포사멸(apoptosis) 조절에 중요한 역할을 하는 단백질로, 세포사멸을 유도하는 Bax와 세포사멸을 억제하는 Bcl-2의 발현 비율은 세포사멸에 대한 지표를 나타낸다. 열처리 검은콩 발효물 유래 조건배지의 신경세포 보호 효과를 평가하기 위해, MPP+를 처리한 SH-SY5Y 세포에 조건배지를 첨가하고 Bax/Bcl-2 발현 비율을 측정하였다(Fig. 8). MPP+ 처리군(+)은 비처리군(-)과 비교하여 Bax/Bcl-2의 발현 비율이 1.77배 증가하는 것으로 나타나, MPP+ 처리에 의해 세포사멸이 촉진되는 것으로 나타났다. MPP+ 단독처리군과 비교하여 HFB-CM 처리군에서 Bax/Bcl-2 비율이 유의하게 감소하는 것으로 나타나(p<0.05), 열처리 검은콩 발효물이 신경세포 사멸을 억제하는데 효과적인 것으로 확인되었다. Bax는 미토콘드리아 막에 구멍을 형성해서 cytochrome C의 방출을 촉진하고 caspase-3의 활성화를 유도하여 세포사멸을 유도하는 반면, Bcl-2는 이러한 작용을 방해하여 세포사멸을 억제한다고 알려졌다(Rosse et al., 1998). Cheon 등(2020)은 MPP+로 처리한 SH-SY5Y 세포에서 Lactobacillus buchneri 사균체 유래 CM 처리에 의해 Bax/Bcl-2의 발현 비율이 유의적으로 감소되었다고 보고한 바 있다.
본 연구 결과, 열처리 검은콩 발효물은 산화적 스트레스로부터 SH-SY5Y 세포의 생존율을 높이고, MPP+로부터 유도된 염증과 세포사멸을 억제하며, BDNF의 발현을 증가시키는 등 뇌신경세포 보호 및 인지능 개선 활성을 나타내어 퇴행성 뇌질환의 예방 및 개선을 위한 건강기능식품 소재로 활용 가능할 것으로 판단된다. 열처리 검은콩 발효물의 신경세포 보호 효과는 열처리에 의해 용출된 exopolysaccharide와 peptidoglycan 등의 사균체 성분, 검은콩의 생리활성 성분인 anthocyanin과 isoflavone, 검은콩 발효 대사산물, 장세포 대사산물들이 복합적으로 효과를 나타냈을 것으로 사료되며, 이에 대한 추가적인 메커니즘 연구가 필요할 것으로 생각된다.
요 약
김치로부터 선별된 L. plantarum SMF470과 L. plantarum SMF796 균주를 스타터로 이용하여 사균체를 함유한 열처리 검은콩 발효물을 제조한 후 항산화 활성 및 신경보호 효과를 평가하였다. 열처리 검은콩 추출물 배지와 비교하여 열처리 검은콩 발효물은 유의적으로 높은 DPPH 라디칼 소거능, ABTS 라디칼 소거능, 총 폴리페놀 함량을 나타내었다(p<0.05). 열처리 검은콩 발효물을 장세포(HT-29)에 배양하여 얻은 조건배지(HFB-CM)를 이용하여 장뇌축을 통한 신경보호 효과를 평가하였다. HFB-CM은 H2O2에 의해 산화적 스트레스가 유도된 SH-SY5Y 신경세포에 대해 높은 보호 효과를 나타내었다. 이와 더불어 HFB-CM은 MPP+에 의해 유도된 독성으로부터 SH-SY5Y 세포를 보호하는 것으로 나타났다. SMF470-CM과 SMF796-CM은 MPP+로 처리된 SH-SY5Y 세포에서 BDNF의 발현을 유의하게 증가시켰으며, 염증성 매개인자인 iNOS와 COX-2의 발현을 억제하였으며, 세포사멸 지표인 Bax/Bcl-2 비율을 유의하게 억제하였다(p<0.05). 본 연구결과 사균체를 함유한 열처리 검은콩 발효물은 항산화 활성, 뇌신경세포 보호 및 인지능 개선 활성을 나타내어 퇴행성 뇌질환의 예방 및 개선을 위한 잠재적인 건강기능성 소재로 활용 가능할 것으로 사료된다.