
서 론
유산균은 그람 양성, 통성 혐기성의 간균 혹은 구균 형태의 세균으로 에너지원으로 당을 이용하며, 발효과정을 통해 젖산을 생성하는 것으로 보고되어 있다(Pot et al., 2014). 또한 생육과정에서 유기산, 과산화수소 및 박테리오신 등의 항균물질을 생산하는 것으로 알려져 있으며, 프로바이오틱스(probiotics)의 형태로 활용할 경우, 장내 세균총을 조절하여 숙주의 면역력을 증진시키고, 장 건강을 개선하는 효과가 있어 건강기능성 식품 산업에서 널리 활용되어 왔다(Shokryazdan et al., 2014).
치아우식증은 구강 감염성 질병의 일종으로 일반적으로 Streptococcus mutans가 생산하는 생물막이 주요 원인이 되어 유발된다 (Forssten et al., 2010). S. mutans는 식품에 포함되어 있는 포도당 및 자당을 이용하여 세포 외 다당류(exopolysaccharide)를 생산하며, 세포 외 다당류를 포함한 S. mutans의 대사산물은 치아 표면의 생물막(biofilm) 형성 초기단계에 관여하여 결과적으로 치태를 형성하게 된다(Zisu et al., 2003). 생물막의 일종인 치태는 항생제나 항균제가 치태 내부로 침투하지 못하게 차단할 뿐만 아니라, 유기산을 생산하는 다양한 스트렙토코커스(Streptococcus)속 구강 박테리아의 생장에 도움을 주어 치태 내 산도를 낮춰서 치아우식을 가속화하는 역할을 한다 (Marsh et al., 2015). 이러한 과정에 따라 치태 내에 축적된 S. mutans를 포함한 병원성 세균들은 치아우식을 유발할 뿐 아니라, 구강 내 상처를 통해 체내로 유입될 경우, 심내막염 및 심혈관 질환 등 전신감염의 원인이 된다 (Nakano et al., 2006). 이에 따라, 구강환경을 개선하여 S. mutans에 의한 치아우식증 및 관련질환을 극복하기 위한 치료와 연구가 진행되고 있으며(Zhang et al., 2020), 특히 유산균 및 유산균 대사산물이 구강 건강 개선 효과가 우수하다는 연구결과를 바탕으로 이를 활용한 구강 기능성 제품들이 개발되고 있는 추세이다 (Mann et al., 2021).
본 연구에서는 전통 발효식품인 김치로부터 유산균을 분리하여, 유산균의 대사물질이 S. mutans의 생육 및 생물막 형성을 억제하는지 평가하였으며, 본 연구에서 분리한 유산균 THYJ-15의 S. mutans에 의한 치아우식증 예방 및 구강 건강을 개선 효과를 가진 기능성 제품 소재로서 개발 가능성을 평가하고자 하였다.
재료 및 방법
연구에 사용한 지시균주 Streptococcus mutans KACC 16833T는 국립농업과학원 농업미생물은행(Korean Agricultural Culture Collection, KACC)에서 분양 받아 사용하였다.
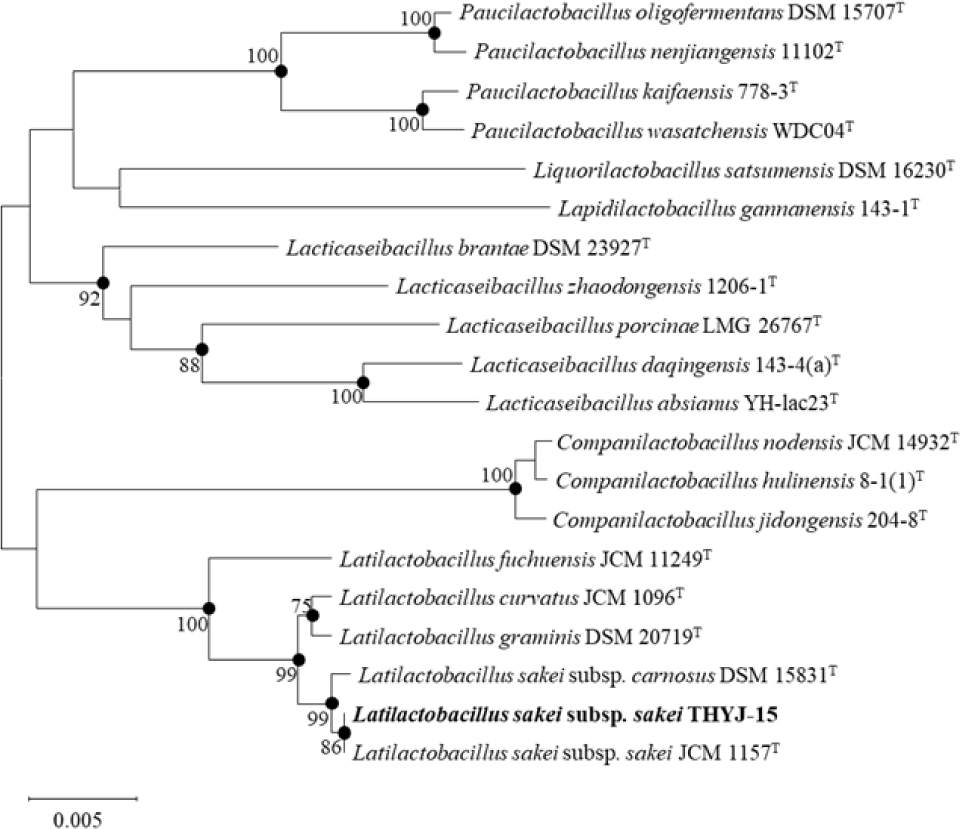
실험에 사용한 김치시료는 멸균 생리식염수에 희석한 후, 유산균 감별 배지인 Bromo Cresol Purple(BCP) plate count agar(E- MB31, EIKEN chemical, Japan)에 획선도말하여 30℃에서 48시간 배양하였다. 이후 젖산(lactic acid)을 생산하여 colony 주위에 노란 환을 생성한 균을 유산균으로 잠정 판단하고 선발하였다 (Yuki et al., 1999). 분리한 균주의 생화학적 특성을 확인하기 위해 API 50 CHL kit(50300, BioMérieux, France) 및 API 20 ZYM kit(25200, BioMérieux, France)를 이용하여 당 이용성 및 효소의 활성을 확인하였으며, 이후 16S rRNA 염기 서열을 분석(BIOFACT, Korea)하여 최종 동정하였다. 분석한 염기서열은 EZBioCloud(www.ezbiocloud.net)의 16S database tool을 기반으로 표준 균주의 염기서열과 비교하여 동정하였으며, MEGA-X 프로그램의 neighbor-joining 및 maximum-parsimony 방법을 사용하여 근연관계 및 계통수(phylogenetic tree)를 확인하였다 (Ventura et al., 2003).
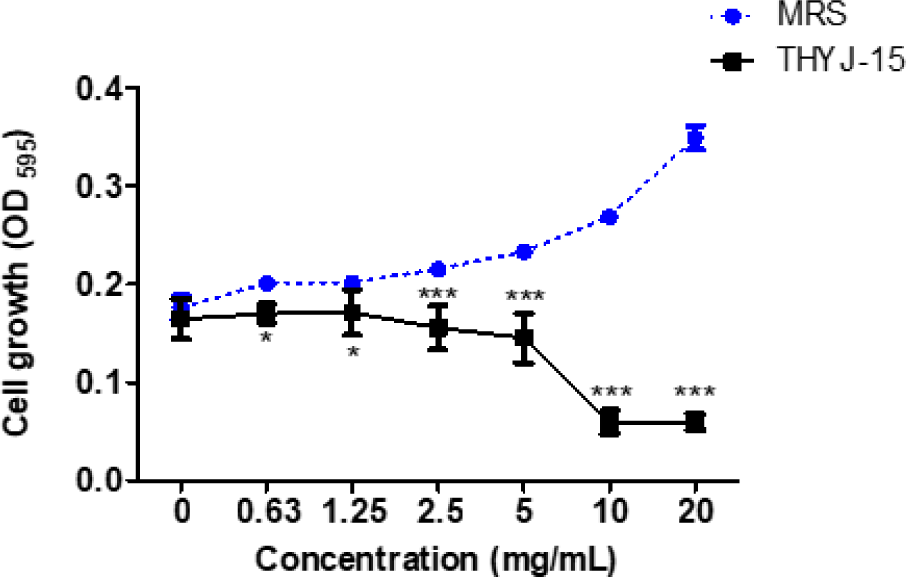
실험은 Wiegand 등(2008)의 액체배지 희석법(broth dilution method)을 참고하여 진행하였다. MRS broth에 종균 배양한 THYJ-15균주를 MRS broth에 1%(v/v) 접종하여 30℃에서 48시간 배양 후, 4,000 rpm에서 15분간 원심분리하여 상등액만을 취해 0.2 µm membrane filter(SN25P020SS, Hyundai micro, South Korea)로 여과하였다. 이후 여과액을 감압 농축한 것을 항균실험의 시료로 사용하였다. S. mutans KACC 16833T는 0.3% yeast extract를 첨가한 tryptic soy broth(211825, Difco, USA)에 1%(v/v)로 접종하여 37℃에서 24시간 배양한 후, 106 CFU/mL로 희석하여 실험에 사용하였다. 이후 96-well microplate(30096, SPL life sciences, South Korea)에 S. mutans KACC 16833T 배양액과 시료를 동량 분주하여 37℃에서 24시간 배양하였다. 이후 균주의 농도 별 생육정도를 확인하기 위하여 ELISA microplate reader(Filter Max F5, Molecular Devices, USA)를 사용하여 595 nm에서 흡광도를 측정하였으며, 음성대조군으로는 시료와 같은 농도로 갑압 농축한 MRS broth를 사용하였다. MIC 및 MBC 값의 확인을 위하여 액체배지 희석법에 따라 항균실험을 실시한 균주 배양액을 0.3% yeast extract를 첨가한 TSB agar plate에 획선도말하여 37℃에서 24시간 배양하여 colony의 생성 여부를 확인하였으며, colony 형성 양상에 따라 최소저해농도(MIC)와 최소살균농도 (MBC)를 설정하였다.
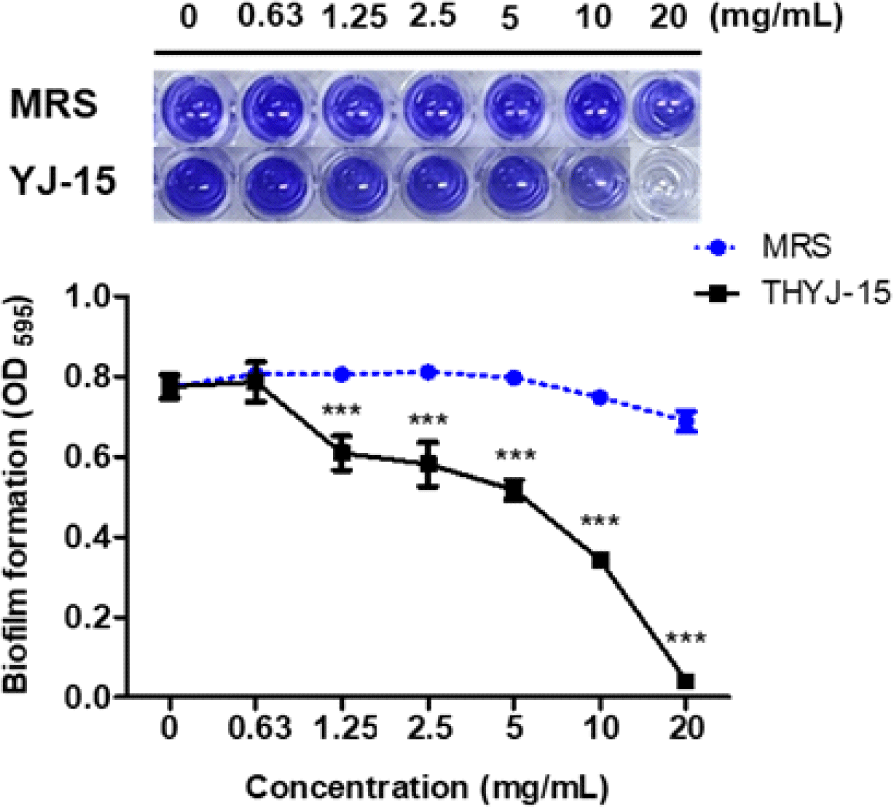
생물막 억제 효과의 확인은 Kouki 등 (2020)의 방법을 참고하여 진행하였다. 먼저 시료를 96-well microplate에 2배수 연속 희석법으로 희석하여 각 well에 100 μL씩 분주한 후, 0.3% yeast extract와 1% sucrose를 첨가한 TSB broth 배지에 2×106CFU/mL의 농도로 희석한 S. mutans를 동량 첨가하였다. 이후 37℃에서 24시간 동안 배양하였으며, 배양 후 각 well을 멸균된 PBS로 2회 세척하여 건조하고, 0.01% 크리스탈바이올렛(crystal violet solution)을 각 well에 첨가하여 15분간 염색하였다. 염색 후 멸균된 PBS로 2회 세척하여 비특이적 염색을 제거한 후 건조하였다. 건조된 well에 각각 33% 아세트산(acetic acid)을 첨가하여 염색된 생물막을 용해시켰으며, ELISA microplate reader를 사용하여 595 nm에서 흡광도를 측정하였다. 실험의 음성대조군으로는 시료와 같은 농도로 감압농축한 MRS broth를 사용하였다.
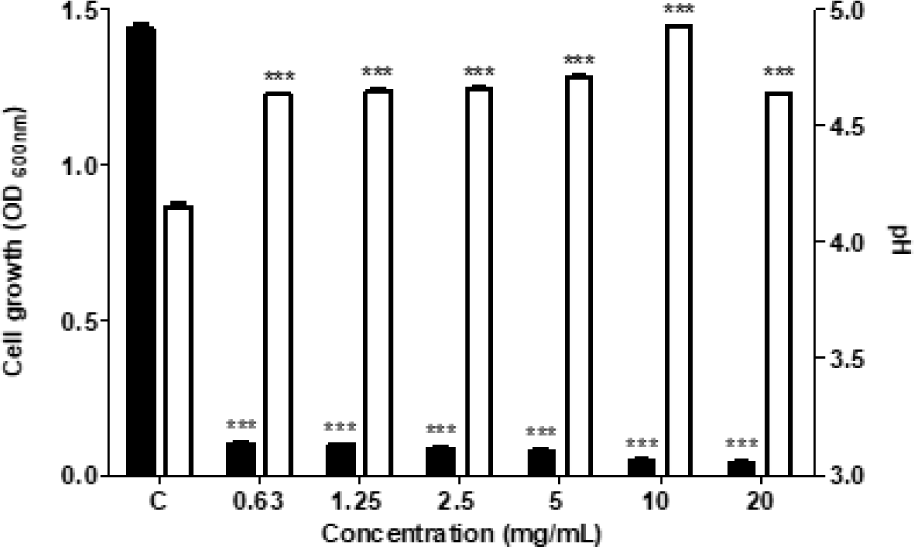
유기산 억제실험을 통해 THYJ-15 균주가 S. mutans의 산 생성을 억제하는지에 대하여 확인하였다. 0.3% yeast extract와 1% sucrose를 첨가한 TSB broth에 시료를 첨가한 후 2배 연속 희석법을 진행하였다. 이후 배지에 S. mutans 16833T 균액을 분주하여 균수를 106 CFU/mL로 조정한 후 37℃에서 24시간 배양하였다. 이후 pH meter(Seven Compact Duo S213, Mettler Toledo, Switzerland)를 이용하여 S. mutans가 배양된 배지의 pH를 측정하였으며, 동시에 UV-Vis Spectrophotometer (OPTIZEN POP, Mecasys, South Korea)를 사용하여 600 nm에서 흡광도를 측정하였다.
S. mutans의 세포 외 다당류를 생성하는 효소인 GTFs의 억제를 확인하기 위해 GTFs의 생성 억제 평가를 실시하였으며, 실험 방법은 Russell 등 (1987)의 방법을 참고하였다. 37℃에서 24시간 배양한 S. mutans KACC 16833T를 4,000 rpm에서 15분 동안 원심분리하고, 상등액의 pH를 7.0으로 보정하였다. 보정 후 0.02% sodium azide을 첨가하여 GTFs coenzyme solution을 준비하였다. 이후 GTFs coenzyme solution과 시료의 혼합액에 2.0% sucrose를 첨가하여 37℃에서 24시간 반응시킨 후 UV-Vis Spectrophotometer를 사용하여 600 nm에서 흡광도를 측정하였다.
결과 및 고찰
본 실험에서 분리한 균주의 16S rRNA 유전자 염기서열 분석 결과, THYJ-15는 표준균주 중 L.sakei subsp. sakei JCM 1157T와 가장 높은 유사도(99.72%)를 가진 것으로 확인되어 L.sakei subsp. sakei THYJ-15로 명명하였다 (Fig. 1). API 20 ZYM kit를 이용한 THYJ-15의 효소 이용 여부 확인 결과, THYJ-15는 Leucine arylamidase를 포함한 7종의 효소의 이용을 확인할 수 있었으며 (Table 1), API 50 CHL kit를 이용한 THYJ-15의 당 이용성 검정 결과, THYJ-15는 L-arabinose를 포함한 15종의 당을 이용할 수 있음을 확인할 수 있었다 (Table 2).
L. sakei subsp. sakei THYJ-15의 S. mutans KACC 16833T에 대한 항균활성 확인 결과, MRS와 THYJ-15를 0.63, 1.25, 2.5, 5, 10 및 20 mg/mL의 농도로 처리하였을 때, MRS 처리군 대비 THYJ-15에서 S. mutans의 생육이 각각 6.53, 15.16, 27.70, 37.62, 77.76 및 82.88% 억제되는 것을 확인할 수 있었다 (Fig. 2). 획선 도말법을 이용하여 THYJ-15의 S. mutans에 대한 최소저해농도(MIC)와 최소살균농도(MBC)를 확인한 결과, 최소저해농도는 10 mg/mL이며, 최소살균농도는 20 mg/mL인 것으로 확인되었다. 항균실험결과, THYJ-15 배양액이 농도의존적으로 S. mutans의 생육을 억제시키는 것으로 확인되었으므로 THYJ-15가 생산하는 대사산물에는 S. mutans의 생장을 억제하는 물질이 포함되어 있어 치아우식증 치료의 기반 물질로서 연구될 잠재력이 있을 것으로 판단된다.
L. sakei subsp. sakei THYJ-15의 S. mutans에 대한 생물막 형성 억제 효과를 확인 결과, THYJ-15 배양액은 농도 의존적으로 S. mutans의 생물막 형성을 억제하는 것으로 확인되었다. 특히 음성대조군인 MRS broth와 비교하였을 때 10 mg/mL 및 20 mg/mL에서 생물막 형성을 54.17% 및 94.15% 더 억제하는 등, 1.25, 2.5, 5, 10 및 20 mg/mL 대부분의 농도에서 유의적으로 생물막 형성을 억제하는 것을 확인할 수 있었다 (Fig. 3). Shemesh 등(2007)의 연구에 따르면 포도당 및 당류는 S. mutans의 생물막 형성을 증가시킨다고 보고되고 있다. 시료 내 포함되어 있는 MRS broth 또한 많은 양의 당류가 포함되어 있으므로 시료의 농도가 높아지면 생물막 형성이 촉진되는 현상이 나타날 수 있다. 그러나 THYJ-15의 경우, 시료의 농도가 증가함의 동시에 시료 내 MRS broth의 농도 또한 증가함에도 불구하고, S. mutans의 생물막 형성을 효과적으로 억제하는 것을 확인할 수 있었다. 이는 THYJ-15의 대사산물이 S. mutans가 생산하는 생물막의 형성을 강력히 억제시키는 효과가 있는 것으로 판단된다.
L. sakei subsp. sakei THYJ-15의 S. mutans에 대한 유기산 생성 억제 효과 확인 결과, THYJ-15 배양액은 0.63, 1.25, 2.5, 5, 10 및 20 mg/mL의 모든 농도에서 샘플 무처리군에 비해 유의하게 S. mutans의 생육 및 유기산 생성을 억제하는 것으로 확인되었다 (Fig. 4). S. mutans는 자당 대사과정에서 많은 양의 유기산을 생성하여 치아우식증을 유발하는 것으로 잘 알려져 있다. Gu 등(2023)의 연구에 따르면 Lactobacillus pentosus MJM60383 배양액이 S. mutans의 자당 대사를 억제하여 치아우식증을 예방하는 것으로 보고되어 있다. 본 연구에서의 THYJ-15 배양액 또한 S. mutans의 자당 대사과정을 저해하여 S. mutans의 생육과 유기산 생성을 억제하는 것으로 판단된다.
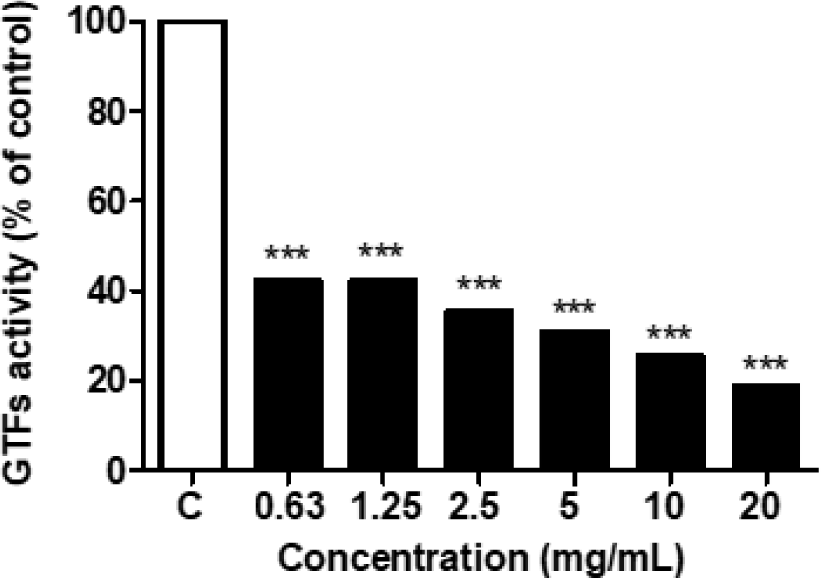
L. sakei subsp. sakei THYJ-15의 S. mutans에 대한 glucosyltransferase(GTFs) 억제 효과 확인 결과, THYJ-15 배양액은 시작 농도인 0.63 mg/mL에서부터 유의한 GTFs의 억제를 확인할 수 있었으며, 0.63, 1.25, 2.5, 5, 10 및 20 mg/mL에서 각각 57.80, 57.79, 64.56, 69.09, 74.41 및 81.17%의 억제율을 확인할 수 있었다(Fig. 5). Lim 등(2020)의 연구에 따르면 Lactobacillus plantarum 200661의 배양액은 S. mutans의 GTFs를 47.03 ± 2.57% 저해하여 세포 외 다당류의 주요 구성성분인 수불용성 글루칸의 형성을 억제한다고 보고되어 있다. 상기 선행 연구결과와 비교하였을 때 THYJ-15 배양액은 20 mg/mL에서 GTFs를 최대 81.17% 억제하여 Lactobacillus plantarum 200661의 배양액보다 높은 GTFs의 억제를 확인할 수 있었다. 이에 따라 THYJ-15의 배양액은 단순하게 S. mutans의 생육을 억제하는 작용과 더불어 생물막의 근본적인 원인인 GTFs를 저해하여, S. mutans의 생물막 형성을 효과적으로 억제하는 것으로 판단된다.
요약
본 연구에서는 김치에서 분리한 유산균인 THYJ-15를 16S rRNA 염기서열 분석법을 사용하여 L. sakei subsp. sakei THYJ-15로 동정하였으며, 균주의 생화학적 특성 및 항치아우식 효과를 확인하였다. 또한 THYJ-15의 항치아우식 효과를 확인하기 위해 치아우식증 유발균 S. mutans KACC 16833T에 대한 항균효과, 생물막 형성 억제 효과, 유기산 생성 억제 효과 및 GTFs 생성 억제 효과를 확인하였다. THYJ-15 배양액은 S. mutans에 대하여 1.25 mg/mL의 농도에서부터 유의한 항균효과를 가지며, 20 mg/mL의 농도에서는 음성대조군 MRS broth에 비하여 82.88% 이상의 생육 억제를 보이는 것을 확인할 수 있었다. S. mutans에 대한 THYJ-15의 MIC 및 MBC 값을 측정 결과, MIC 및 MBC 값은 각각 10 mg/mL 와 20 mg/mL로 확인되었다. THYJ-15 배양액은 S. mutans에 대해 1.25 mg/mL의 농도에서부터 유의한 생물막 형성 억제 효과를 가지며, 10 mg/mL의 THYJ-15 배양액은 MRS broth와 비교하여 54.17% 이상의 생물막 형성 억제를 보이는 것을 확인할 수 있었다. THYJ-15의 S. mutans에 대한 유기산 형성 억제능은 실험 결과 0.63 mg/mL부터 20 mg/mL까지 모든 농도에서 유의한 억제 효과를 확인할 수 있었으며, 가장 높은 유기산 형성 억제능은 10 mg/mL에서 확인할 수 있었다. THYJ-15의 GTFs 억제능 또한 0.63 mg/mL부터 20 mg/mL까지 모든 농도에서 유의한 효과를 확인할 수 있었으며, 배양액의 최고 농도인 20 mg/mL에서는 81.17%의 억제율을 확인할 수 있었다. 결론적으로 김치에서 분리한 유산균 L. sakei subsp. sakei THYJ-15는 치아우식증 유발 균주 S. mutans에 대한 강한 항균효과와 GTFs의 활성 억제 효과를 기반으로 생물막의 형성을 억제하고, 부가적으로 유기산 생성 억제 효과를 가져 치아우식증에 대한 치료제 및 기능성소재로 개발가능성을 지니고 있음이 증명되었다. 향후 구강건강 개선 핵심 기반소재로서 개발하기 위해 THYJ-15가 생산하는 대사산물을 분석하고, S. mutans를 포함한 다른 구강병원균들에 대한 항균 효과 및 생물막 형성 억제 효과 평가 등의 추가연구가 필요할 것으로 사료된다.